杨炜迪++高婷++朱建宁+++王锦++王川

摘要:以宁夏中部干旱带引进的适生牧草品种(710、708、376、859)和蒙古冰草根际微生物为研究对象,采用 Biolog-Eco方法研究了5种牧草的根际微生物群落功能的多样性。结果表明:5种不同牧草根际土壤微生物活性存在差异;利用羧酸类、碳水化合物最多的是牧草376,利用聚合物类、酚类化合物、氨基酸类最多的是708,利用胺类最多的是蒙古冰草;5个品种主要利用碳源类型为碳水化合物、羧酸类和氨基酸类。丰富度指数与优势度指数结果相似,为859≥蒙古冰草≥710>708>376;多样性和均匀度指数结果一致,为蒙古冰草>859>708>376>710;主成分分析将5种品种分为3个功能类群,376和708为一类,859和蒙古冰草为同一类,710为一类;载荷矩阵表明5中牧草品种在羧酸和氨基酸利用上差异最大。
关键词:Biolog-Eco;根际土壤;功能多样性;牧草
中图分类号: S154.3文献标志码: A文章编号:1002-1302(2016)02-0360-04
收稿日期:2015-08-13
项目基金:宁夏农林科学院科技创新先导资金(编号:NKYJ-2014-11);宁夏自然科学基金(编号:NZ1073)。
作者简介:杨炜迪(1983—),女,河南偃师人,硕士,助理研究员,研究方向为牧草育种。E-mail:12345ywol@163.com。
通信作者:高婷,硕士,研究员,研究方向为牧草新品种选育。E-mail:ecogt9@163.com。根际是指植物根周围数毫米的区域[1],此区域中根分泌物为微生物提供重要的营养和能量物质并影响着根际微生物的种类和繁殖,而根际微生物的代谢作用又直接促进或抑制根的营养吸收和生长,从而影响根际土壤中的物质转化[2]。Biolog-Eco 法是在微生物利用31种碳底物时发生氧化还原反应,通过颜色变化情况来反映微生物碳源利活性的方法[3-4]。该技术广泛应用在评价和比较不同土壤类型[5]、相同土壤不同植物物种[6]、不同植被的根际土壤微生物群落功能多样性[7]。本研究以4种美国引进牧草和蒙古冰草的根际微生物为研究对象,应用 Biolog-Eco方法,对不同牧草品种根际土壤微生物代谢功能多样性进行比较研究,旨在探讨不同牧草品种根际微生物代谢功能多样性之间的差异,研究结果可为深入探讨植物多样性和土壤微生物多样性之间的关系奠定基础。
1材料与方法
1.1试验地概况
试验地设在宁夏吴忠市盐池县高沙窝的天然草场。草地类型为荒漠草原,植被稀疏且草层低矮,主要植被类型为蒙古冰草(Agropyron mongolicum Keng.)、糙隐子草(Cleistagenes squarrosa)、胡枝子(Lespedeza bicolor Turcz.)、老瓜头(Cynanchum komarovii Al.)、甘草(Glycyrrhiza)、细叶骆驼篷(Peganum nigellastrum)、银灰旋花(Convolvulus ammannii Desr.)。盖度为17%~30%,种类组成贫乏。该区属于典型的大陆性气候,春季干旱多风,夏季炎热,年均降水量260.75 mm,主要集中在7—9月,月平均温度最高月为6月(23.5 ℃)、7月(24 ℃)和8月(20.5 ℃),土壤为风沙土,土壤有机质 1.4%~7.5%,pH值9.08~9.22。
1.2牧草品种
本试验选取4个引进美国品种(710、708、376、859)和本地优势品种蒙古冰草开展研究。710品种耐寒性、耐旱、耐高温能力强;708粗蛋白质含量高、干草产量高;376青绿期长、适口性好;859株高大、鲜草产量高;蒙古冰草植株高大,生长速度快。
1.3播种
将美国引进的牧草品种,随机排列,小区面积为25 m2,行距25 cm,对试验地隔带深翻后,于2011年4月28日至5月1日播种,深翻比例介于30%~80%,每品种重复3 次。取样时引进牧草品种生长年限为4年。
1.4土壤采集
6月下旬在盐池县高沙窝天然草场对5种牧草品种按五点取样法选取牧草植株,每点每品种取3~5株,重复3次。将根系区域土样用铁铲挖出,抖落根系外围土,取紧贴于根表周围的土样。将土样装入无菌纸袋,标记,立即带回实验室,研磨过1 mm筛后于4 ℃冰箱保存。
1.5牧草根际土壤微生物群落功能多样性研究
Biolog-Eco分析:本研究采用有31 种碳源的生态板(规格为EcoPlate,由Biolog 公司生产,华粤企业集团有限公司提供)分析微生物群落的代谢特征。每品种土样均匀混合后按照四分法称取10 g,加入90 mL无菌生理盐水稀释,在摇床里振摇30 min,静止沉淀3~5 min,然后进行100倍稀释,以每孔150 μL稀释液加入微孔板中,将制备好的菌悬液倒入无菌移液槽中,使用八孔移液器将其接种于微平板的96孔中。接种好的微平板放到铺有6层纱布的塑料饭盒中,为防止微平板鉴定孔中的菌悬液挥发,纱布保持一定的湿度。塑料饭盒用保鲜膜包裹,保鲜膜上用注射针头扎若干个小眼,以保证微生物的培养所需要的氧气,将微平板避光培养。ECO生态板放到30 ℃恒温培养。分别于24、48、72、96、120、168 h时读数,测定波长为590 nm。
2结果与分析
2.1不同牧草品种AWCD变化及利用碳源分析
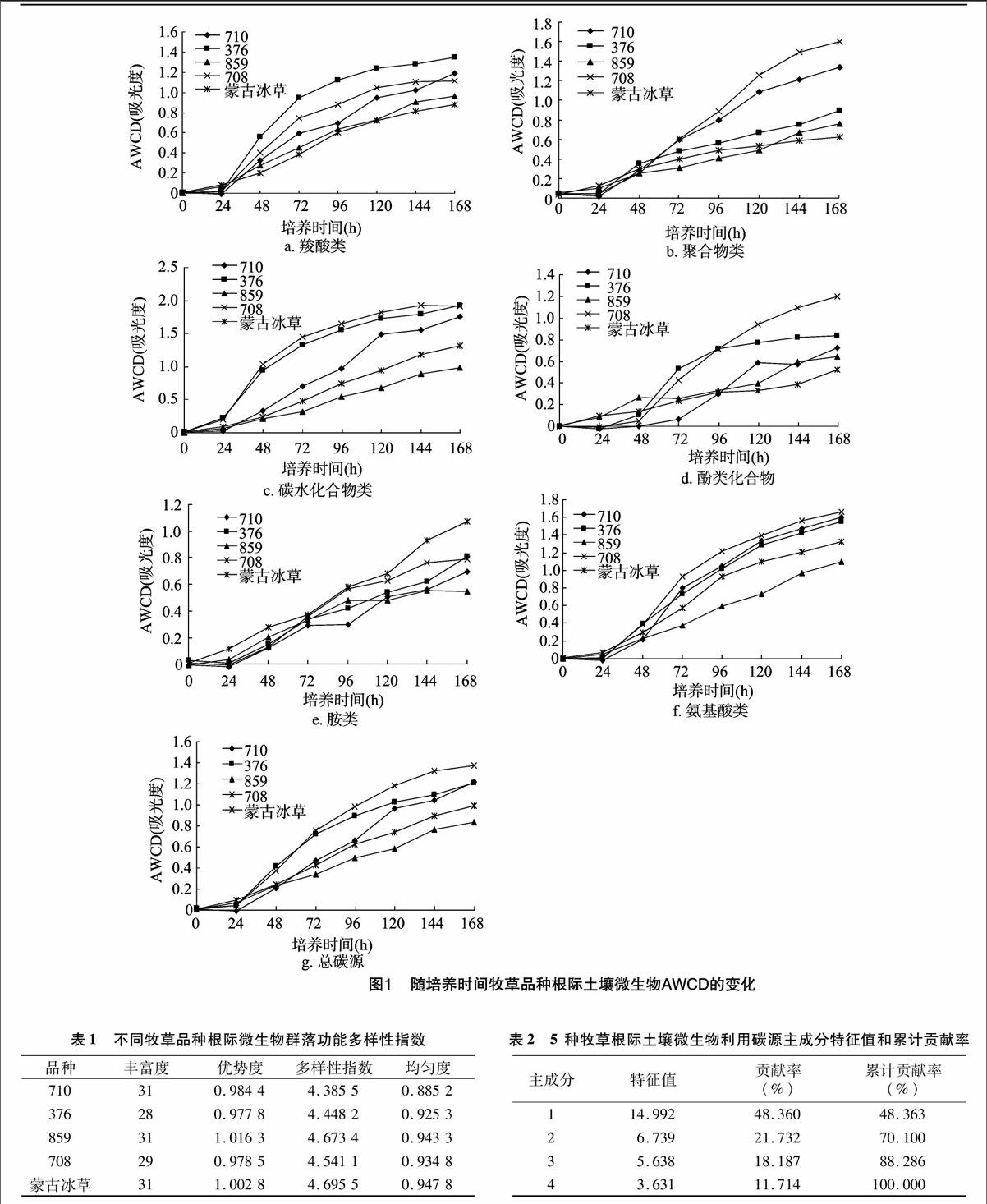
从图 1 可知,5种牧草品种根际微生物群落利用碳源量随培养时间的延长逐渐增加。每种牧草根际土壤的 AWCD 值在 24 h 内变化不明显,表明该时期根际微生物活性较低,碳源利用很少;在 24 h 之后显着增加,碳源利用明显。在整个培养过程中,各品种根际微生物 AWCD 值存在差异,培养168 h 时,总碳源AWCD值为708>710>376>蒙古冰草>859。羧酸类AWCD值为376>710>708>859>蒙古冰草,聚合物类AWCD值为708>710>376>859>蒙古冰草,碳水化合物AWCD值为376>708>710>蒙古冰草>859,酚类AWCD值为708>376>710>859>蒙古冰草,胺类AWCD值为蒙古冰草>376>708>710>859,氨基酸类AWCD值为708>710>376>蒙古冰草>859,5种牧草主要利用碳源为碳水化合物、氨基酸和羧酸类。
2.2牧草品种根际微生物群落功能多样性分析
本研究采用丰富度、优势度、多样性指数和均匀度来比较5个牧草品种的土壤根际微生物群落利用31种碳源的多样性。丰富度指数表示被利用的碳源种类多少,优势度指数可反映出土壤群落中最常见的物种,多样性指数反映微生物群落物种的变化度和差异度,均匀度为群落物种丰富度与均一性的度量。由表1可知,丰富度与优势度指数为859≥蒙古冰草≥710>708>376;多样性指数与均匀度分析结果一致,蒙古冰草>859>708>376>710,说明859、蒙古冰草根际微生物群落中某些优势菌生长旺盛。以上研究结果表明蒙古冰草根际微生物群落种类最多且较均匀。
2.3不同牧草品种根际微生物群落功能多样性主成分分析
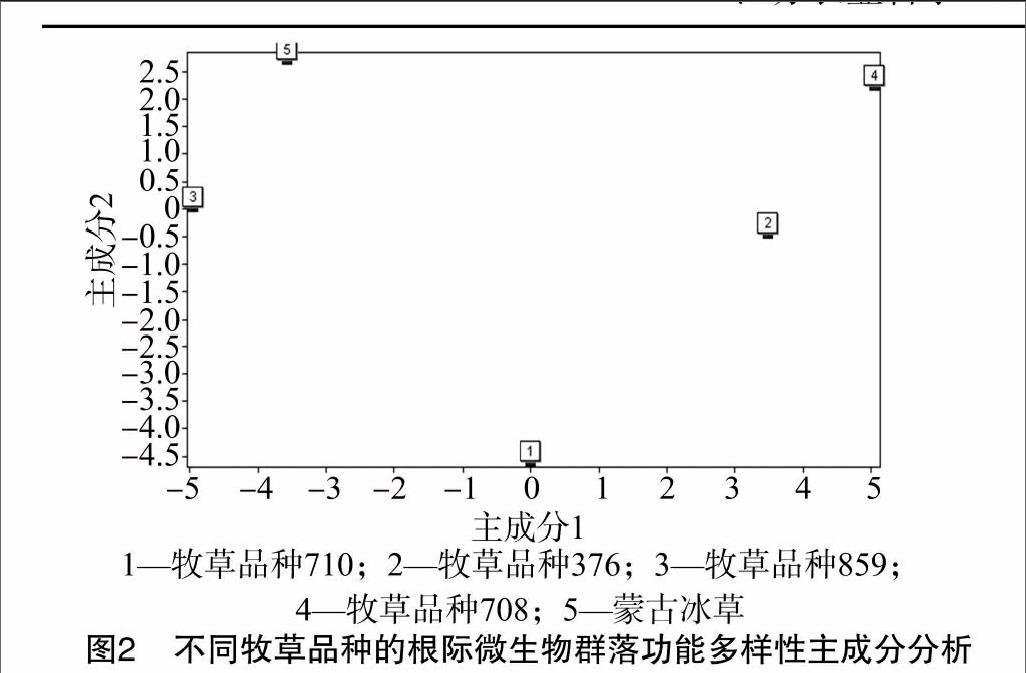
采用主成分分析比较5个品种根际微生物群落功能多样性(表2)。总变异的100%为前4个主成分,即包含全部变异信息。总变异的48.36%、21.73%分别来自第1、第2主成分,故第1、第2主成分为变异主要来源。利用这2个主成分将微生物群落代谢功能划分成3个功能类群(图2),376和708土壤根际微生物群落为同一类,710品种土壤根际微生物群落为同一类,859和蒙古冰草土壤根际微生物群落为同一类,空间差异较大。
因子载荷矩阵表明(表3),因子1主要由x(30)、x(15)、x(25)起分异作用,对应的碳源为D-苹果酸、L-丝氨酸、L-磷酸葡萄糖;因子2主要由x(19)、x(18)、x(27)起分异作用,对应的碳源为L-苏氨酸、γ-羟丁酸、苯乙胺。5种牧草根际微生物群落在以上6种碳源利用上差异最大,主要为羧酸类和氨基酸类。
3结论与讨论
本试验通过Biolog-Eco微平板方法证实了5种不同牧草的根际土壤微生物在代谢活性、不同碳源利用、微生物多样性指数等方面均有自身特征并存在差异,说明不同牧草品种在退化土壤生态环境的修复中提供不同作用,导致这种差异的主要因素是牧草品种、植物残体、根系分泌物等方面[11]。毕江涛等研究表明,荒漠草原 5 种植被类型土壤微生物群落代谢活性和利用碳源种类都存在差异[12];安韶山等对宁南山区9种典型植物根际微生物研究表明微生物活性(AWCD)、微生物多样性指数和均匀度指数均存在明显差异[8]。利用Biolog-Eco[9-10]、16S rRNA 分析[13]、PCR-DGGE[14]等方法开展的研究均显示不同植被类型、同一植物不同品种或不同基因型间以及同一基因型不同发育时期的根际土壤微生物多样性都有所不同。
本试验结果表明,不同牧草根际土壤的AWCD在24 h内变化不大,随培养时间增加,其利用碳源量也不断增加,与诸多文献结果[8,15-16]一致,708、376、710的根际土壤微生物代谢活性较强,蒙古冰草和859根际土壤微生物代谢活性较弱。各品种在各类碳源利用上存在差异,利用羧酸类、碳水化合物最多的是376,利用聚合物类、酚类化合物、氨基酸类最多的是708,利用胺类最多的是蒙古冰草,5个牧草品种主要利用碳源类型为碳水化合物、羧酸类和氨基酸类,与吴则焰等研究的武夷山各植被类型土壤微生物利用主要碳源类型[7]一致。另外,郑华等研究表明4种森林类型土壤微生物群落比较偏好、利用率较高的3类碳源是糖类、羧酸类和氨基酸类[5]。根际微生物群落功能多样性分析表明,5种牧草的丰富度和优势度指数结果相似,多样性和均匀度指数结果一致,向泽宇等研究表明,土壤微生物群落多样性与均匀度指数也表现出相似规律[17]。蒙古冰草根际微生物群落种类最多且较均匀,原因可能是蒙古冰草与其他牧草品种生长年限不同[18-19],且蒙古冰草为原生植被,而其他4个品种为人工种植的牧草品
种[20];859和蒙古冰草根际微生物群落中某些优势菌生长旺盛,利用碳源类型较其他3个品种多。
5个品种根际微生物主成分分析将5个品种土壤根际微生物群落碳代谢功能分为3类,376和708为同一类,710品种为一类,859和蒙古冰草为同一类,与各品种碳源代谢活性
和根际微生物群落功能多样性分析结果一致。林青等研究发现新疆富蕴地震断裂带6种次生植物根际土壤微生物碳源利用差异主要体现在糖类、氨基酸类、羧酸类[16]。因子载荷矩阵表明,5种牧草根际微生物群落主要在羧酸和氨基酸的利用上差异最大。
参考文献:
[1]刘芷宇,施卫明. 根际研究方法[M]. 南京:江苏科学技术出版社,1996:308-327.
[2]王茹华,张启发,周宝利,等. 浅析植物根分泌物与根际微生物的相互作用关系[J]. 土壤通报,2007,38(1):167-172.
[3]Garland J L,Mills A L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization[J]. Applied and Environmental Microbiology,1991,57(8):2351-2359.
[4]田雅楠,王红旗. Biolog法在环境微生物功能多样性研究中的应用[J]. 环境科学与技术,2011,34(3):50-57.表3因子载荷矩阵
类别碳源类型因子1因子2因子3因子4x(1)β-甲基-D-葡萄糖苷0.892 5-0.115 70.236 80.365 9x(2)D-半乳糖酸γ-内酯0.004 5-0.704 1-0.172 30.688 9x(3)L-精氨酸0.167 8-0.829 30.376 70.377 1x(4)丙酮酸甲酯0.582 2-0.026 60.760 9-0.285 1x(5)D-木糖/戊醛糖0.831 3-0.550 80.014 20.072 8x(6)D-半乳糖醛酸0.393 4-0.905 9-0.121 4-0.098 9x(7)L-天门冬酰胺0.273 3-0.604 20.429 00.613 3x(8)吐温 400.015 2-0.910 20.074 10.407 3x(9)赤藓糖醇0.274 4-0.350 90.895 3-0.002 2x(10)2-羟基苯甲酸-0.510 50.853 3-0.102 20.028 8x(11)L-苯丙氨酸-0.090 10.688 50.258 60.671 5x(12)吐温 80-0.099 6-0.966 40.236 9-0.004 0x(13)D-甘露醇0.515 0-0.813 20.271 1-0.000 6x(14)4-羟基苯甲酸0.931 7-0.338 60.127 70.030 4x(15)L-丝氨酸0.976 7-0.136 5-0.164 80.011 2x(16)α-环式糊精-0.131 00.232 80.896 60.353 1x(17)N-乙酰-D葡萄糖氨0.911 8-0.271 20.247 70.183 4x(18)γ-羟丁酸-0.183 40.919 4-0.290 7-0.191 4x(19)L-苏氨酸-0.144 70.926 10.347 80.019 0x(20)肝糖0.543 90.148 30.822 40.075 9x(21)D-葡糖胺酸0.509 6-0.160 60.619 00.575 6x(22)衣康酸0.551 7-0.016 9-0.821 00.146 1x(23)甘氨酰-L-谷氨酸0.598 70.401 3-0.037 20.692 2x(24)D-纤维二糖0.826 4-0.486 40.154 80.237 5x(25)1-磷酸葡萄糖0.939 5-0.276 3-0.202 10.005 0x(26)α-丁酮酸0.257 00.477 9-0.832 6-0.110 7x(27)苯乙胺-0.284 60.912 10.137 30.261 2x(28)α-D-乳糖-0.040 4-0.123 00.040 90.990 7x(29)D,L-α-磷酸甘油0.641 5-0.139 40.009 90.754 3x(30)D-苹果酸0.978 1-0.137 8-0.090 3-0.126 7x(31)腐胺0.828 20.252 50.487 90.110 8
[5]郑华,陈法霖,欧阳志云,等. 不同森林土壤微生物群落对Biolog-GN板碳源的利用[J]. 环境科学,2007,28(5):1126-1130.
[6]李世朋,蔡祖聪,杨浩. 不同植被下红壤性质对细菌碳源利用的影响[J]. 应用与环境生物学报,2008,14(6):793-797.
[7]吴则焰,林文雄,陈志芳,等. 武夷山国家自然保护区不同植被类型土壤微生物群落特征[J]. 应用生态学报,2013,24(8):2301-2309.
[8]安韶山,李国辉,陈利顶. 宁南山区典型植物根际与非根际土壤微生物功能多样性[J]. 生态学报,2011,31(18):5225-5234.
[9]Grayston S J,Wang S Q,Campbell C D,et al. Selective influence of plant species on microbial diversity in the rhizosphere[J]. Soil Biology & Biochemistry,1998,30(3):369-378.
[10]张萌萌,敖红,张景云,等. 建植年限对紫花苜蓿根际土壤微生物群落功能多样性的影响[J]. 草业科学,2014,31(5):787-796.
[11]Kowalchuk G A,Buma D S,De Boer W,et al. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms[J]. Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology,2002,81(1/2/3/4):509-520.[12]毕江涛,贺达汉,沙月霞,等. 荒漠草原不同植被类型土壤微生物群落功能多样性[J]. 干旱地区农业研究,2009,27(5):149-155.
[13]Kaiser O,Puhler A,Selbitschka W. Phylogenetic analysis of microbial diversity in the rhizoplane of oilseed rape (Brassica napus cv. Westar) employing cultivation-dependent and cultivation-independent approaches[J]. Microbial Ecology,2001,42(2):136-149.
[14]Smalla K,Wieland G,Buchner A,et al. Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis:Plant-dependent enrichment and seasonal shifts revealed[J]. Applied and Environmental Microbiology,2001,67(10):4742-4751.
[15]张海涵,唐明,陈辉,等. 黄土高原5种造林树种菌根根际土壤微生物群落多样性研究[J]. 北京林业大学学报,2008,30(3):85-90.
[16]林青,曾军,马晶,等. 新疆地震断裂带次生植物根际土壤微生物碳源利用[J]. 应用生态学报,2011,22(9):2297-2302.
[17]向泽宇,张莉,张全发,等. 青海不同林分类型土壤养分与微生物功能多样性[J]. 林业科学,2014,50(4):22-31.
[18]曹成有,姚金冬,韩晓姝,等. 科尔沁沙地小叶锦鸡儿固沙群落土壤微生物功能多样性[J]. 应用生态学报,2011,22(9):2309-2315.
[19]王阳,王奇赞. 种植年限对大棚蔬菜地土壤微生物群落结构多样性的影响[J]. 浙江农业学报,2013,25(3):567-576.
[20]熊毅,郭彦萃,张志. 原始红松林与人工林土壤微生物多样性分析[J]. 浙江农业学报,2014,26(1):159-164.单爱琴,肖洁,杨秀婧. 四氯化碳与四氯乙烯复合污染对土壤呼吸率的影响[J]. 江苏农业科学,2016,44(2):364-367.



